Weather forecasts only look out a few days to weeks. Two new research studies describe the increasing accuracy of specialized scientific models in forecasting changes in the ocean up to a year in advance.
The models have increasing value as climate change drives shifts in ocean temperatures and other conditions with new and unexpected outcomes. The changes can have ecological and economic repercussions. For example, warming ocean temperatures increase the overlap between fishing fleets and protected species like whales and sea turtles, which can trigger fishing closures. The research was conducted by scientists at NOAA Fisheries and University of California, Santa Cruz.
 Read the full story here:
Read the full story here:

 E.L. Hazen, 2022. Climate change is “heard” in the ocean depths. Nature Climate Change. DOI: 10.1038/s41558-022-01484-5.
E.L. Hazen, 2022. Climate change is “heard” in the ocean depths. Nature Climate Change. DOI: 10.1038/s41558-022-01484-5. 
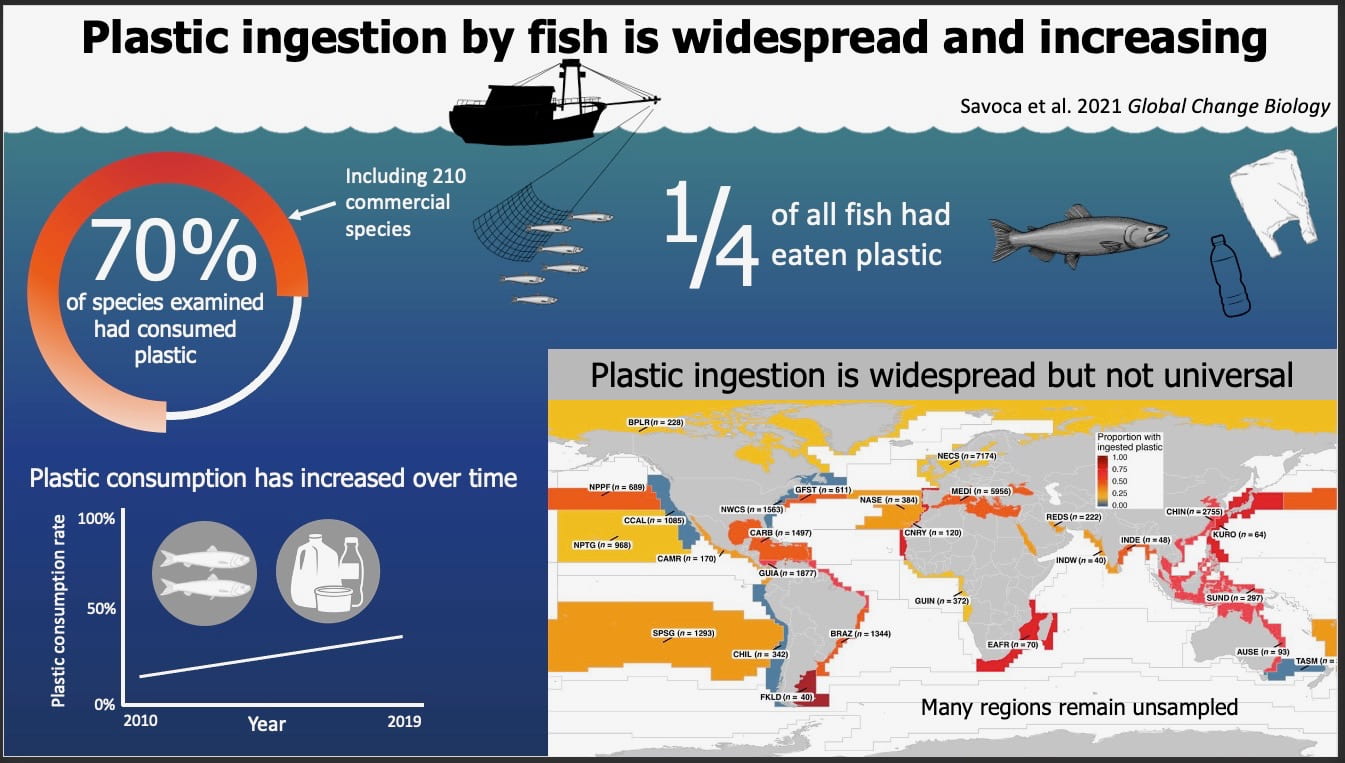
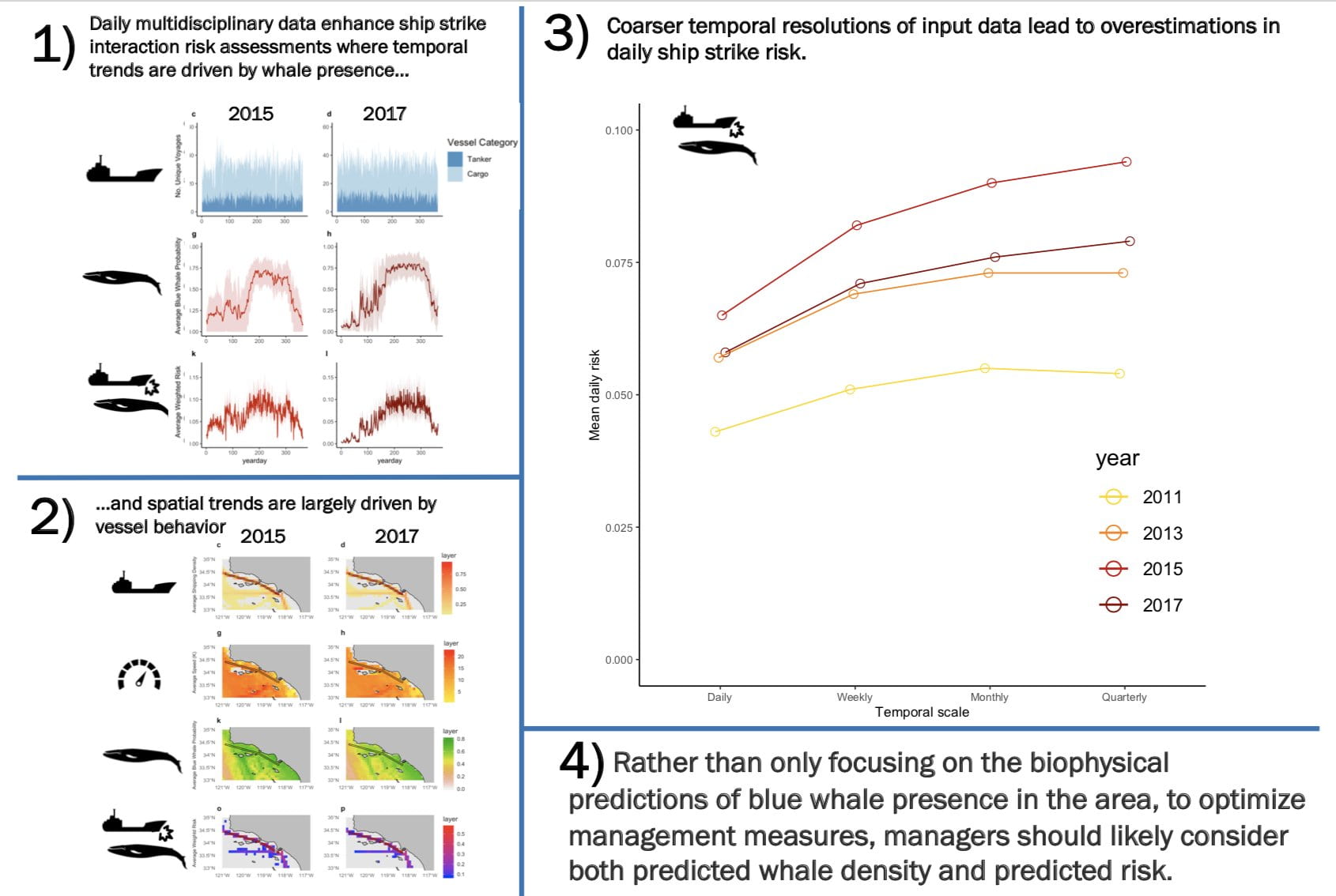
 Infographic (c) Stephanie Brodie
Infographic (c) Stephanie Brodie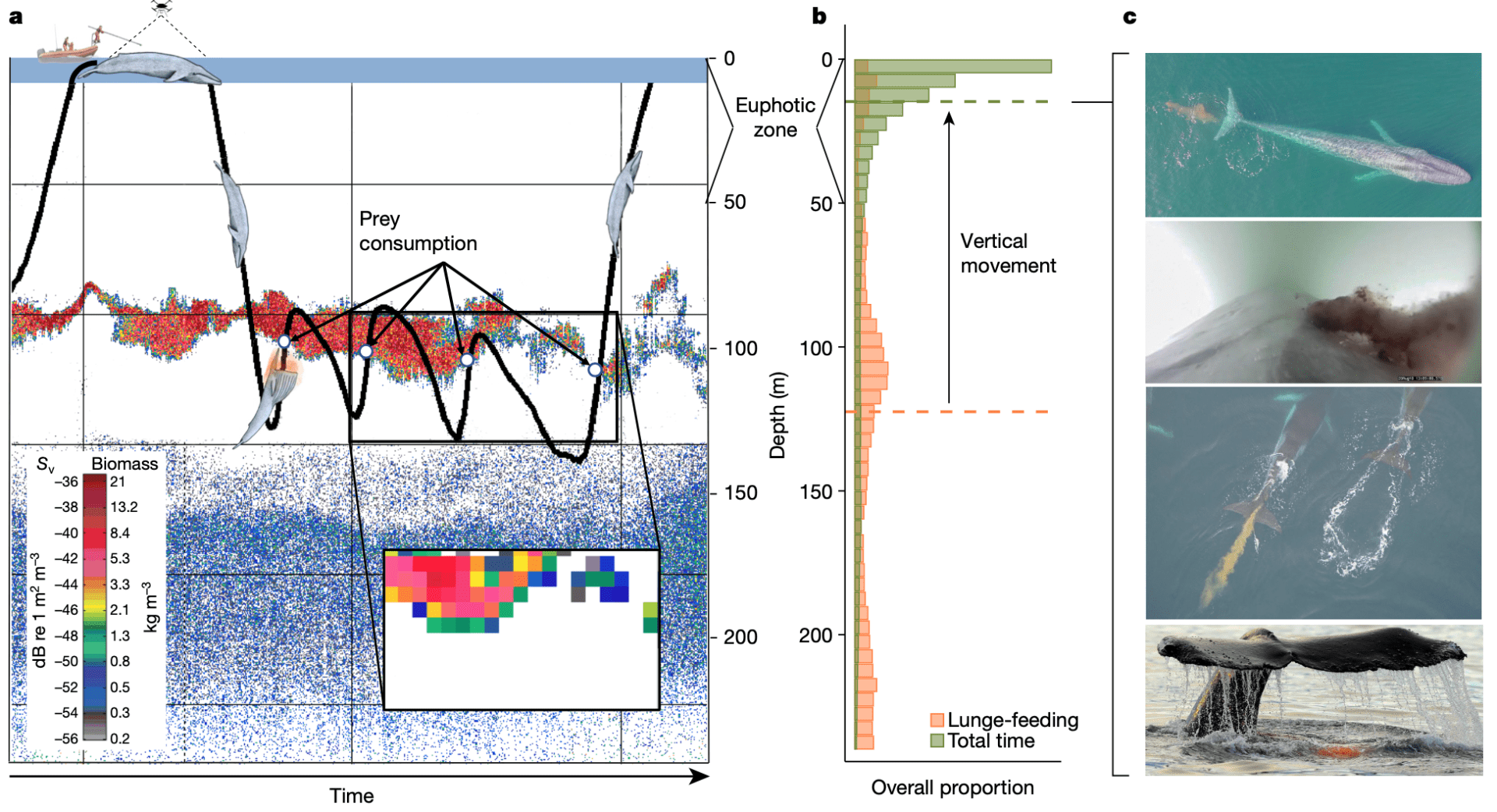 Figure 1. Fieldwork needed to measure whale size, behavior, and the density of the prey they eat. The vertical difference in foraging and feces may redistribute nutrients to keep marine ecosystems flowing.
Figure 1. Fieldwork needed to measure whale size, behavior, and the density of the prey they eat. The vertical difference in foraging and feces may redistribute nutrients to keep marine ecosystems flowing.