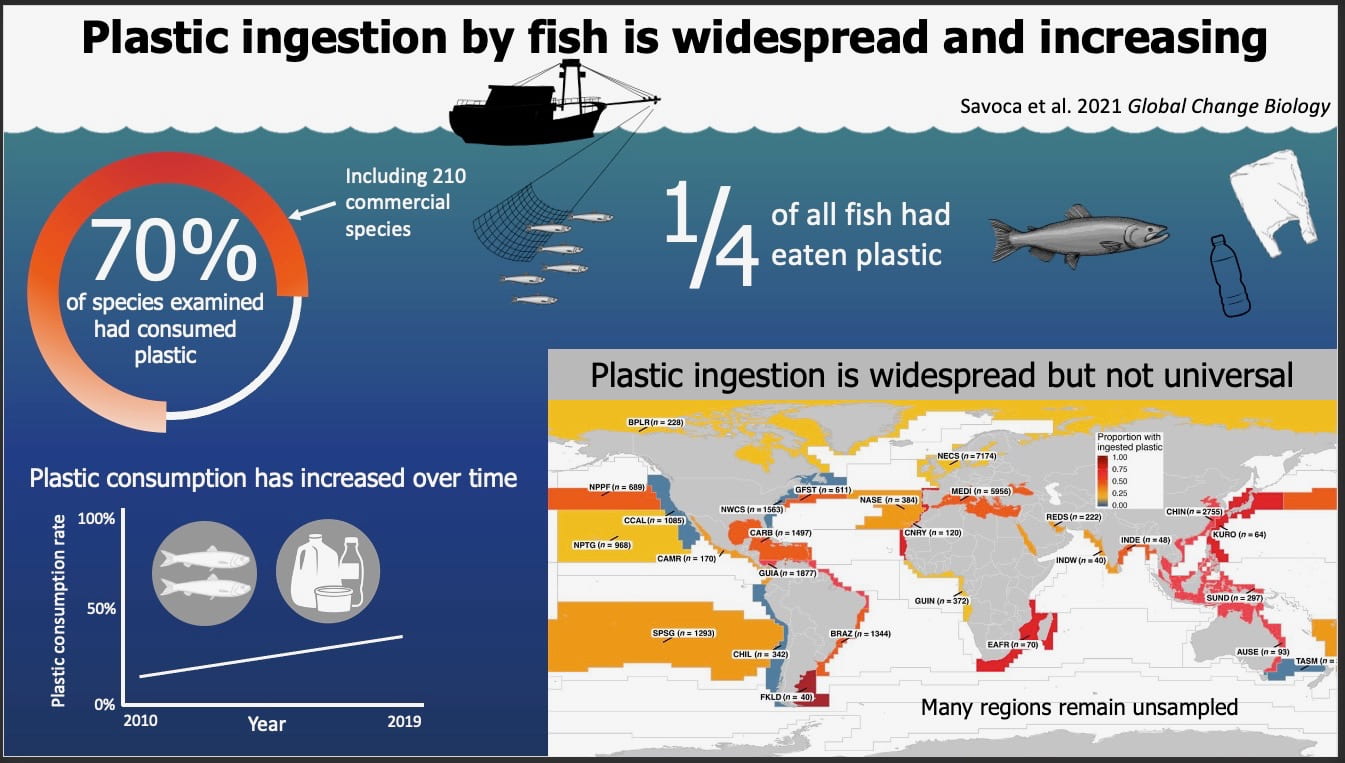
Researchers have developed global forecasts that can provide up to a year’s advance notice of marine heatwaves, sudden and pronounced increases in ocean temperatures that can dramatically affect ocean ecosystems.
“We have seen marine heatwaves cause sudden and pronounced changes in ocean ecosystems around the world, and forecasts can help us anticipate what may be coming,” said lead author Michael Jacox, a research scientist at NOAA Fisheries’ Southwest Fisheries Science Center in Monterey, California, and NOAA’s Physical Sciences Laboratory in Boulder, Colorado.
Marine heatwave forecasts will be available online through NOAA’s Physical Sciences Laboratory. The researchers called the forecasts a “key advance toward improved climate adaptation and resilience for marine-dependent communities around the globe.”
M.G. Jacox, M.A. Alexander, D. Amaya, E. Becker, S.J. Bogard, S. Brodie, E.L. Hazen, M. Pozo Buil, and D. Tommasi, 2022. Global seasonal forecasts of marine heatwaves. Nature 604, 486–490. PDF
Read the abstract of the paper below:


 E.L. Hazen, 2022. Climate change is “heard” in the ocean depths. Nature Climate Change. DOI: 10.1038/s41558-022-01484-5.
E.L. Hazen, 2022. Climate change is “heard” in the ocean depths. Nature Climate Change. DOI: 10.1038/s41558-022-01484-5.  Infographic (c) Stephanie Brodie
Infographic (c) Stephanie Brodie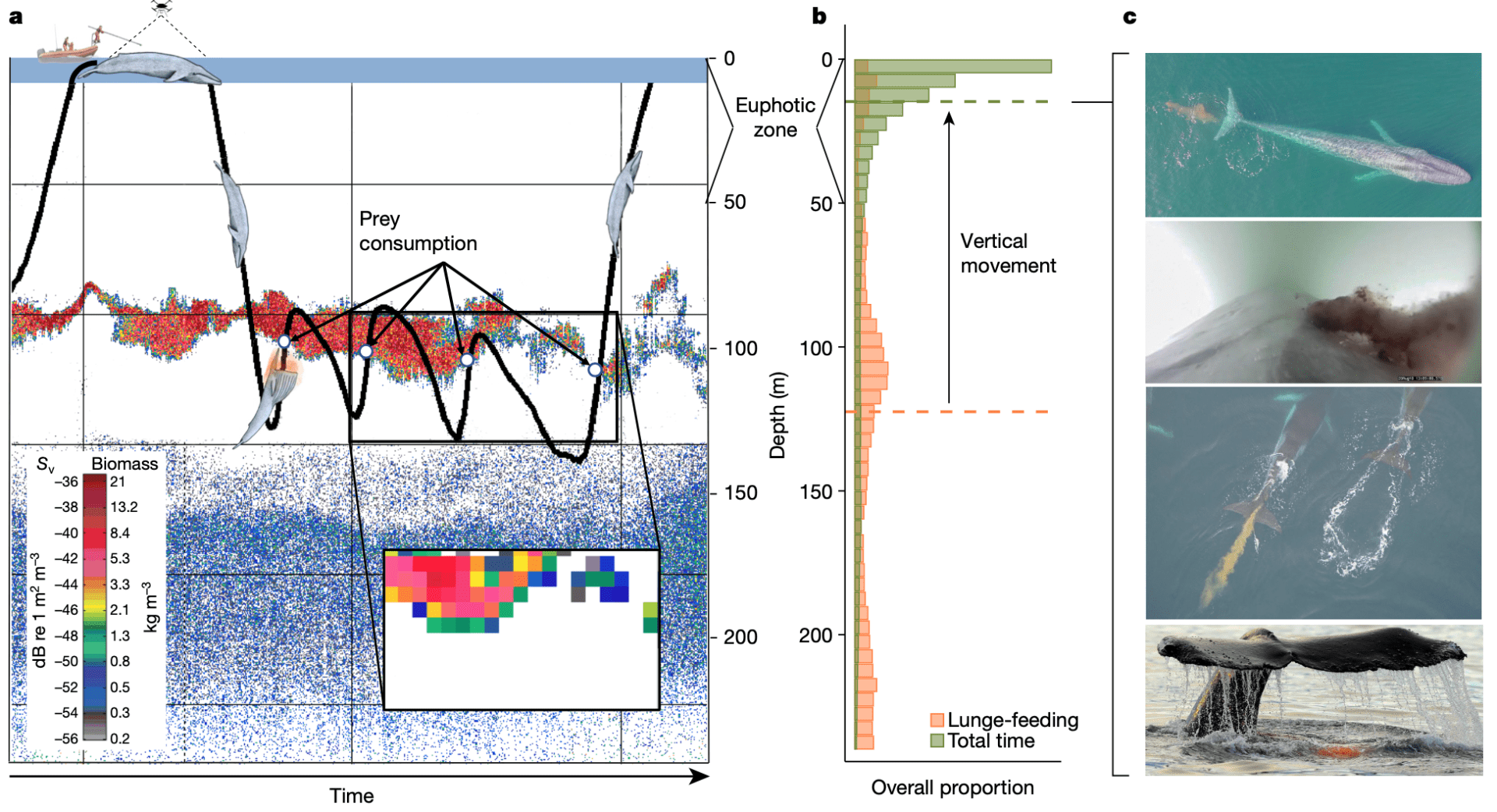 Figure 1. Fieldwork needed to measure whale size, behavior, and the density of the prey they eat. The vertical difference in foraging and feces may redistribute nutrients to keep marine ecosystems flowing.
Figure 1. Fieldwork needed to measure whale size, behavior, and the density of the prey they eat. The vertical difference in foraging and feces may redistribute nutrients to keep marine ecosystems flowing.